����������У��������ص������л���������Ѿ��������йؼ���������ض����źŵ��о���֤��������ļ����о��Ѿ���Խ����Щ��صķ��������ṩ��֤�ݣ�����ͨ���ض�Ƶ�ʵļд��������Ըı���书�ܡ����������֤������Ҫ����Ϊ���������������ֱ����صĻ��ƺͽ����Ǽ���ĸ������������������������ò�ͬ�������յ������Ե����������Ĵ̼��о���������Ϊ����Щ�о�֤���˴����ͼ���֮��������ϵ��������Եĸ���������۵��෴�����ķ�����Trends in Neurosciences��־��(�������ź�siyingyxf��18983979082��ȡԭ�ģ���˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ���)��
�¹ʶ�֪�£�������������ؽ���Ķ���ֱ�ӵ����������������ź�siyingyxf��18983979082��ȡԭ�ļ������������
���Ƴ�40Hz tACS�����ĺ�Ĭ�����ߺ�����ע��Ӱ��
ͨ���Ե�ͼ/�Դ�ͼ�۲쵽�Ĵ��Ի��ָ�����Դ̼�
TMS-EEG���ٴ�Ӧ�ü�չ��
�����ĺ�Ĭ���������Դ̼�
Trends in Neurosciences��������Ϣ�������Դ̼�����
PNAS�������������ϵ����Ӻͼ��ٻ���Ӧ���Ӻͼ�����������е��
����Ƶ���������֪���Ʋ�ͬ�ɷ��е��������
˯�ߡ����Դ̼����ϻ��Ĵ����о�
Nature Medicine����������̼����Ը���ǿ��֢
����̼��������Ͳ���˥�Ϲ������龰�����Ӱ��
The Neuroscientist������TMS��EEG��MRI���������������
ʹ�ô̼��豸����·���ز���Ծ�����������
CURRENT BIOLOGY: �Ⱥͦ����ڹ���������������õ����֤��
BRAIN��TMS-EEG�о������Է�ӦΪ���к���˶��ָ��ṩ���廯����
Ƥ���˶��˷��Բ���������mu������λ�ĵ���
TMS�CEEG���Ϸ������������Ƥ��������̽���еĹ���
ʵʱEEG������TMS������֢���������ǰ��ҶƤ���������ͬ���̼�
��������̼���tACS�������������˹�������Ļָ�
tACS���EEG�о�:������������
��������̼�(tACS)��ʹ���Խ���ͬ���������֪����
1. ���ͼ���
����(������)��������Ⱥ֮���ͬ���������á����䱻��Ϊ��Ҫ������ͻ�����ӵı仯����ͻ��������������Ԫ֮���ͬ��ˮƽ����ˣ��������Ǽ�����̵���Ҫ���֡�����أ�����Ϊ��ͬ��Ƶ��������delta��theta��alpha��beta��gamma��(��������)��ֵ��ע����ǣ�������ЩƵ��(�������Ǹ��������۵�theta��gamma����)�������������йء����⣬��������������йأ���������������֪�����йء���Щ���Ʒ��ض��Ĺ�ϵ�����һ����������������ʵ�ֵĻ�����ʲô��������η����ڼ���?Ŀǰ��������Ϊ���ڲ�ͬ��ʱ��Ϳռ�߶��Ͻ��л����Ļ���͵����㡣��Щ����������Է����ڼ�����ͬ����֪���̣�����������֮һ����ˣ���ʹʱƵ��ģʽ���ղ����������֪���̾�����ϵ����������Ҳ������������Ϊ�����������Ļ�������(����Ϣ��ά�֡���ͨ�źͼ��ʱ�������Ŀ�����)��������ǰ���������Ѿ����������ִ���֪��ѧ��һ����ҪĿ�����ȷ����Щ������ʲô��
����(brain oscillaitons)������Ƶ������ԪȺ�Ľ����Ի���������ڲ�ͬ�Ŀռ�߶��ϲ������Ӿֲ�����λ(����1����)�����Ե�ͼ(intracranial electroencephalography, iEEG��Լ1����)�����Դ�ͼ(magnetencephalography, MEG��1-2����)���ٵ�EEG (Լ������)��
�����ϵ(causality)���������x (���磬��)����y�ı仯(���磬����)����ô��������x��y�������صġ�
�д�(entrainment)������ָ��������õ���ϵͳ�ڸ��Զ�������ʱ���в�ͬ�����ڣ����Ǽٶ�һ����ͬ���ڵĹ��̡�������ѧ�У���������ϵͳ��(i)���ɴ̼���(����˸�̼��������塢�������Ҳ���)��(ii)�ܴ̼�����Ⱥ��
�д��ز�(entrainment echoes)���Լд���խ����Ӧ����д��̼����࣬���ȴ̼����ٳ������������ڡ�
�龰����(episodic memory)�����ض������ļ��䣬ͨ���Ǵ��Դ���ĽǶȣ������ּ����У�ʱ�䡢�ص��������������Ϣ���Ա���ȷ�س��������ڲ����롣
��������(working memory, WM)���ڶ�ʱ����(ͨ���Ǽ�����)��ά�ֺͲ�����Ϣ�Ĺ��̡�WM������(capacity)������������Ŀ(items)��������ֱ�ӷ��ʡ�
Ȼ�����ڽ�ʾ��Щ����֮ǰ��������֪����(������)�ʹ�����֮���ϵ�ı����Ǻ���Ҫ�ġ��������˵���ͼ���֮��Ĺ�ϵ��һ�������ϵ������һ�ָ�����ϵ?
��������������������ϵ����ô���Ǿ�������ϣ����ʾ�����ڼ������ض��Ļ������á�Ȼ������Ҫ����Ҫ��ס���κ���֪����(�����)�������м���DZ�ڵ�ԭ����ˣ��ɹ۲쵽�ļ�����ǿЧӦ���������ڲ�ͬ����֪����(���磬��ǿ��ע�����������Ի�����εļӹ�)����ÿһ�����������λ��ڸ��ֻ����ļ�����ƺ����Ǽٶ�����Ӧ������ԡ����仰˵���������ƿ�����ijһ�ض���֪���ܵij�������������ǿ��ܲ��Ǹù��ܵı�Ҫ��������Ϊ��Ҳ����ͨ����������ʵ�֡�
������������ÿ���ͨ��������ʵ�������ԣ����������ֵ����Ƿ�Լ������Ϊ������Ӱ�졣�ĵ��ƿ���ͨ���д�ʵ��(��1)���д��ı�����Ȼ���������Ӷ�̽����Ȼ��������״̬���������ԡ���Ҫ�����ַ������շ��ǵ��ͽ����Ի�ķ�������������֮�����ֻ�������ܴ̼�������Ȼ���ڵ�(����ݿ��Ʒ�)����Ҳ���ܲ�����֪��������п���������ʽ���֣���Ը���Ȥ�ļ�����Ϊ�����������Ӱ�졣����������У�����ͨ�����ֲ�ͬ�Ĵ̼��������й㷺�ļд���(i)�о��д���(ii)����/�żд���(iii)�д���д�����Щ�д�����Խ��Խ������ڻ�����ѧ�о�������������ͼ���Ļ������õľ������⣬�Լ�ּ�ڸ��ƽ����ͷǽ������Լ��书�ܵ�Ӧ���о����ڽ��������о��У����ǻع�����Щ������ص��ע���ڹ�������(working memory, WM)���龰���������мд��Լ�����ֵĿ���Ӱ�졣���ڶԼ�����ر��ע������������ǰ��һЩ����������ͬ�����������ϸ���۵ģ�����������£���������С�Ŀ�����һ����Ҫ�����⣬��1�г��˱������۵Ĺؼ��о��ĸ������Լ����Ǹ��Ե�������С��
��1 ���д�
��һ���ܴ̼��������ԪȺ���üд��̼�����λʱ���д��ͻᷢ��(ͼIA)���д��̼���Ⱥ��������Ӱ�죺
(i) ����Խ��Խ�����Ԫ��д��̼���λ���룬�ź�ǿ��(����)���ӣ�
(ii) Ⱥ����д��̼���λ���롣
������Ҫ���ǣ��д�����˲�䷢���ģ�������Ҫʱ��ģ���ȡ���ڴ̼�ǿ�ȡ������������Ⱥ�ڼд������б��뾭����һϵ��״̬���������Ⱥ���ڻ���״̬����Ԫ�ʶ�ͬ�������ֻ���״̬���Բ�ȡ������ʽ��������ֹ״̬��(���磬����alpha��)��ʵ����Ƶ�״̬(���磬�Ӿ���դ����gamma����ռ䵼������theta)����Ҫ��һ���ǣ������д������ڼд���ʼ֮ǰ��Ⱥ�����ǿɼ���(��ƫ��1/f��)����ˣ����ʵ���Ƶ��ȱ������ϵͳ�Ͳ��ܱ��д����ڶ����ǽ����Σ���Ⱥ��ʼ���ܼд��̼���������Σ���Ⱥ���λ��ʼ��д��̼�����λ�ƶ�������������ȫ�д�״̬��������Ⱥ������ȵؼд���������Σ���Ⱥ�ͼд��̼�֮�����λ�ͺ�Ϊ�㡣�ڼд��̼���ֹ����Ⱥ�����ָ�������״̬�������д�������
�м��ֲ����д��ķ�����һ�ַ����Dz����ڼд��������ܼ�����Ĺ��ʺ���λ��Ȼ�������ǵ��̼�αӰ����ӵ��¼���ط�Ӧ(�¼���ص�λ��event-related potentials, ERPs)���Ⲣ����������ġ���һ���棬�д��ز������Լд��������ֱ�ӵ���������������ù�������ʱ����ͿĨ(smearing)��(����еĻ�)�����ơ���һ�ֲ����д��ķ�������ν��Arnold�ࣨ��ŵ����ͷ(Ӣ���ø���tongues)��һ����ѧ����,��˵����������һ�ֹ���Ĺ���������(ͼIB)������Ҫ���Ʋ�ͬƵ���¼д�ǿ����̼�ǿ�ȵĹ�ϵͼ��Arnold��������һ��������ǿ��ֻ��д�����Ƶ�� (������)�����Ŵ̼�ǿ�ȵ����ӣ�ϵͳ�в����ڵ�Ƶ��Ҳ�ᱻ�˹��յ���
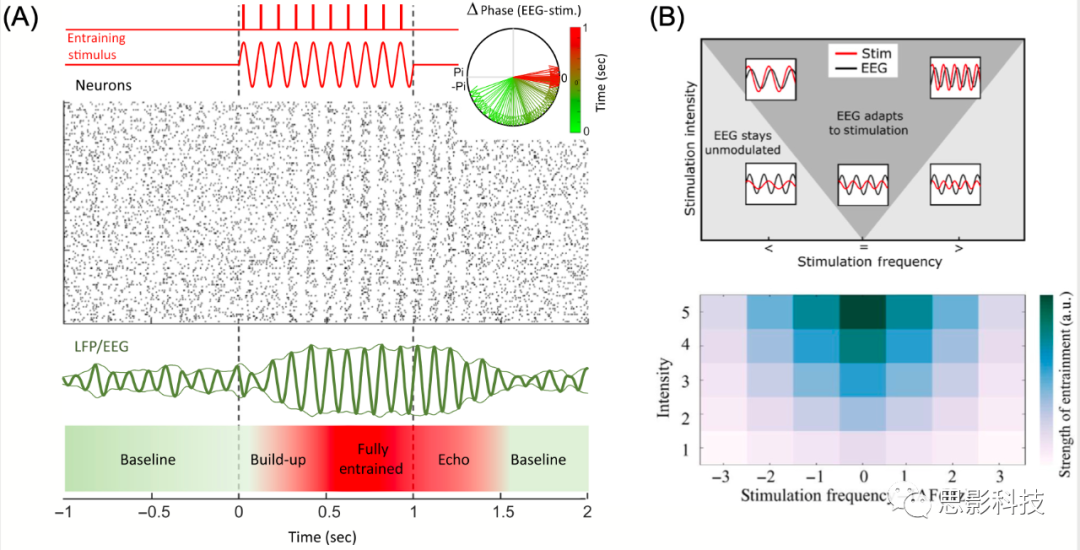

ͼI �д��Ļ���ԭ���Ͳ���������
(A)��ԪȺͨ�������̼�(���Ҳ�)������̼��д���դ��ͼ��ʾ����Ⱥ��ģ���ֵ����ֲ�����λ(local field potential, LFP)/�Ե�ͼ(electroencephalography, EEG)��ʾ��Ⱥ��ˮƽ�Ļ�����Ϸ���ʾ�˼д��̼�����ԪȺ�֮�����λ���죬��ɫ��ʾ�Ӽд���ʼ(��ɫ)������(��ɫ)��ʱ�䡣
(B) Arnold�ࡣ�̼�ǿ��(y��)���ڲ�Ƶ��(x��)ͼ���ʹ̼�ǿ��ֻ���ڴ̼�Ƶ�����ڲ�Ƶ��(internal frequency, IAF)ƥ��ʱ�Ż�����д���

��1 �漰�Լ������Ӱ��ļд��о�����
2. �о��д�
ͨ���о��д�(��2)���յ������Ե��ڼ������ܵ��뷨�������µġ�Williams���֣��ڱ�������У�������˸�����ȣ��Լ��������Ͽ�Ŀ���Ƶ����ȣ���10 Hz(��alpha)����˸����£������ߵ�ʶ���������ӡ�ʹ�����������Դ̼�Ҳ�õ������Ƶķ��֣���betaƵ�ʷ�Χ��˫�����Ĵ̼�����������ӡ���Ȥ���ǣ���������ĺ�Ĭ��С��ģ�͵��о���������gammaƵ��(40 Hz)����������Ԫ������beta-�������߿��ˮƽ�����ֶ�beta-�������߿������������������(���Ŵ�ѧ)�̼��ͷ������Ըо���˸�ж����ڣ�������о��д�ȷʵ��Ӱ�캣���Ļ(�ʹ��Խṹ)��Becher������ͬ���ܹ�֤ʵ���о�����̼���Ӱ�������ڲ��Ҷ�����ͬ��ˮƽ��������Щ���֣�ʹ�øо�����̼������Ƽӹ������̼�������֮���ͬ���̶��ƺ��ǿ��еģ����ⷴ����Ӧ�û�Ӱ����䡣������˵������������£����ϴ̼�������Ԫ��(����������Ƶ)���Ա��ֱ���ڣ�����������һ���������ͬ���ģ�����һ����������첽�ġ������ͬ��ȷʵ�ڵ���ͻ�������Է��淢�������ã���ô���ֲ���Ӧ�û�Ӱ����䡣
��2 ���ֲ�ͬ�ļд�����
��ͨ���о��̼��д�
��Ԫ���ϣ��ر�������о�����ģʽ���Ӧ�ĸо���������ѭ�ⲿ���ִ̼���ʱ�䶯��ѧ����ˣ����ְ������ɽ��ɳɷֵĴ̼�������Ч�շ������������������С�ֵ��ע����ǣ���Щ���ɲ���ֻ�ڸо����������Ҳ�ᴫ�ݵ������������纣����
��ͨ��tES/rTMS�д�
����̼�(Transcranial electric stimulation, tES)��һ��ͨ���缫��������ʩ�ӵ�ͷƤ�ϵļ�����ʵ���ϣ�tES������һϵ�еĴ̼������������ɳɷֵľ�̬����������ֱ����̼�(transcranial direct current stimulation, tDCS)����������0�����ľ�������̼�(transcranial alternating current stimulation, tACS)���������ߵ���ϣ����в�������ֵ��0֮�����ظ����Ŵ̼�(Repetitive transcranial magnetic stimulation, rTMS)ͨ��ʩ�ӳ�����ʱ�����壬������֯���յ�����(ͼ2)��rTMS������������۽��ԣ������ض������°����ض��������������������ڲ��Ծֲ����������������������ġ�˫���̼�(������rTMS��Ȧ�Ĵ̼�)����������ˮƽ�϶�����������ԡ����֮�£�tES���нϵ͵Ŀռ�ֱ��ʣ���ͨ������Ϊһ��ȱ�㣬�������о���Ŀ�꣬ʵ���Ͽ���������ģ����統������Ҫ�ڹ㷺������ˮƽ��̽�����������ʱ��rTMS��tES֮���һ����֮ͬ������rTMS�����շ�������λ(�����ϵ�λ)����tESͨ��ֻӰ��ֲ�����λ(�����µ�λ)�����ڼд����������һ����Ҫ��������ΪrTMSԭ���������յ����˹��������tESֻ��ͨ������������ڽ��еĻ����ˣ�tESЧӦ���ܸ����Ҳ������ֱ�ӵIJ��Լд���
��ͨ�������Ե�̼��д�
�����̼�(Deep brain stimulation, DBS)�ǶԽ����д���¼��/��̼�������������(����ﻼ��)������֯����ֱ�ӵ�̼��������ٴ�����ʹ�����л������ø��ִ̼�����(��������ⲿ�̼�������ڲ�״̬��λ�á������Ƶ�ʺ�ʱ��)����¼�ʹ̼��������������ͬ�ĸ���Ȥ�����̼��İе����Ƥ���������Ľṹ���纣����Ҳ�������������ʹ�����ڲ��Ҷͨ·����������Ƥ�㡢�ڲ�������������Ҷ�ͻ���������ʺ�(ͼ3)�������Դ̼���ȷ����������ΪӰ����δ��ȫ�˽⡣ֱ�Ӵ̼�Ƥ���Ƥ����������Ϊ�յ����ܴ̼������������ƺ��˷ܵĸ��ӵ���ЧӦ���Լ�ͨ����ͻ���ӵ��ܴ̼������Զ����������ƺ��˷ܵĸ��ӵ���ЧӦ��
��������뷨��Clouter������ͬ�����龰���䷶ʽ�г��ֶ�о������̼��������ַ�ʽ�У�������ס����-��Ƶ��������Щʵ��������������ѧ�о�������ѧ�о�����theta����λ������ʱ����ǿ(long-term potentiation, LTP)�ͳ�ʱ������(long-term depression, LTD)�Ĵ��ڡ����ǿ��Դ���Щ�о����ƶϳ������ڲ�ͬ��(LTP��LTD�յ���) theta��λ��ƫ����Ԫ�ŵ�������ȣ���ͬ��LTP�շ�theta��λ��ƫ����Ԫ�ŵ�����Ӧ�õ��¸��õļ��䡣��һ����ͬ��������ų�Ŀ�ܣ���theta��������������֪����ϵ�������ر������Ӿ��������������������ڣ��ڲ��Ľ��ཫ�����ƣ���������ѵĸо���Ϣ����������Щ����Ļ����ϣ�Clouter������ͬ����֤����������첽���ִ̼���ͬ�������������Ӿ��̼���������õļ���(ͼ1A)����������(1.7 Hz)�Ϳ����(10.5 Hz)��ȣ�ͬ������������첽���������ּ��������ض���theta����(4 Hz)����Ȥ���ǣ����첽�̼�����Ȼ�̼�(��δ���Ƶĵ�Ӱ-������)��ȣ�4 Hz��ͬ���̼��ܴ������õļ��䡣��Щ���ֱ��ظ�����չ��֤����������������У��������Ӿ����������ڵ�һ����ˮƽ�ϸ���д��ij̶ȿ���Ԥ������ļ��䡣��һ����������У������������Ӿ�������theta����Χ�ڵ�ͬ����ǿ���¸��õļ��䡣���Roberts������ͬ���ǵ�һ���о������������д���ѧϰ�Ͳ���֮���theta��������������ļ���ı�������֮����Щ�о�ǿ����theta��ͬ������������������Ҫ�ԣ����������������������γ�������������á�
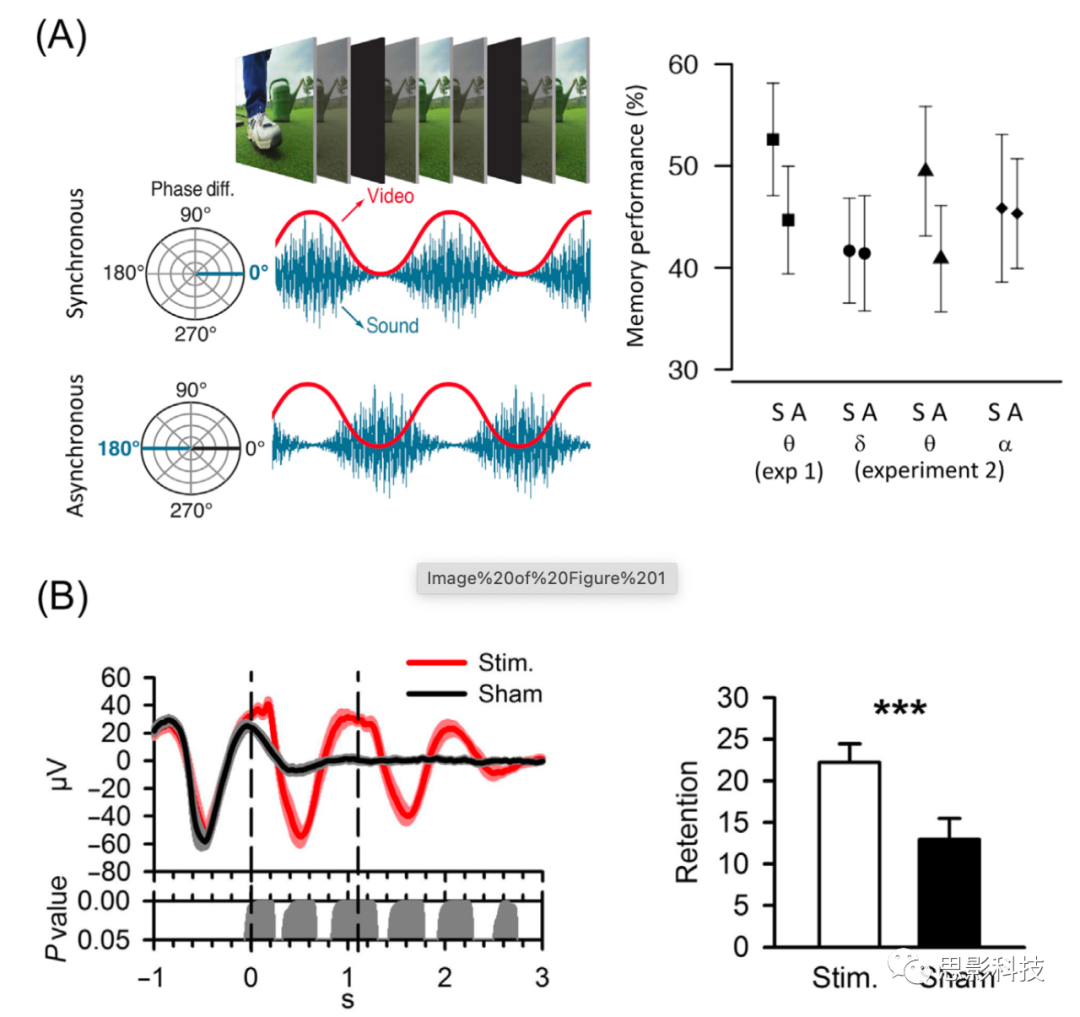
ͼ1����������(A)��˯��(B)�ڼ�ͨ���ĸо��д������ڼ��䡣
(A) ������ƵƬ�β���3�롣��Ƶ(��ɫ)����Ƶ(��ɫ)�ֱ���theta (4 Hz)Ƶ�����Ҳ��½�������/���������ڼ������Σ���Ƶ�����������ƣ�ʹ��Ӧ�Ĵ�������Ҫô��ͬ���(ͬ������)��Ҫô�Dz�ͬ���(�첽����)�����첽�̼�(A)��ȣ�ͬ���̼�(S)������������(��ͼ)���á�����ЧӦ���ض���theta (4 Hz)�ģ�ʹ�ý���(delta, 1.6 Hz)��Ͽ�(alpha, 10.4 Hz)�ļд�Ƶ������á�
(B) ��˯���ڼ�ͨ���ջ��̼����������������̼���λ��ͨ���¼���ص�λ(event-related potentials, ERPs)��������������ʱ����һ�������̼�(��ͼ)����ɫ�ĺۼ���ʾ�̼�������ERP����ɫ�ĺۼ���ʾ�ٴ̼�������ERP(����¼�����û��ʩ�Ӵ̼�)�������ջ��̼������˼��乮��(��ͼ)��
�����������������ѵĴ����в�����Ҳ������˯���в���(���磬ͨ�����������Դ̼�)�����ַ����������ض���˯�ߵ����źţ������������ڼ��乮�̹����е�������á�������˯���ڼ�ļд����ر����������ģ���Ϊ������û����ʶ���̼�������ų����κκ�����ΪЧӦ��������͡�Ngo����ͬ�����ñջ������̼�֤��������Դ�������ͬ���������̼�����ǿ������ͼ��乮��(ͼ1B)����ЩЧӦ�������˺����ὡ��ѧ������˯�б����ơ�Ȼ����˯���ڼ�������̼�ͨ�������������ͷĴ�������ʹ�ú���ȷ��������ֵ��κα仯�Ƿ�����������Ĵ�����ı仯����һ�������ǣ���Щ�о������̼��Լ��乮�̵����̵�ֱ��Ӱ���������ڼ��乮�̵������ļ�Ӹ���(������ж���л��)�ֿ���Ȼ������Щ�о���������˯���ڼ���ĸо��д����������������
3.����(tES)�͵��(rTMS)�д�
�������д��ķ����Ǿ��̼��������羭��̼�(transcranial electric stimulation, tES)���ظ����Ŵ̼�(repetitive transcranial magnetic stimulation, rTMS����2)���ڽ��������о��У����Ǹ��ݰ���Ƶ�ʵļд����з����о���������(delta)��ʼ�����ǻ��ر��ע�ڼٶ���Щ�����ǻ�Ծ������µ������о�(�����ߴ̼��о�)������ע������Ϊ�⣬���ṩ����ѧ���ݱ�����ȷʵ�Ѿ����д����о�(��1)��
��������֪������˯����������乮���������ϵ�ĵ�һ��֤��(�ڼд��о��ı�����)����Marshall���ˣ�������˯�߲�������ע��0.75 Hz�ĵ�ǿ�ȵ���[ͬʱ���о�ֱ����̼�(transcranial direct current stimulation, tDCS);��2]�����ִ̼�������������������ʾ�˼д���֤�ݣ�Ҳ�յ��˸��õļ�����֣��������������ڼ��乮���е�������á���Ȥ���ǣ�������ʱ�յ����Լ���Ҳ�����Ƶ�����Ӱ�죬����ǿ��theta��beta�Ե�ͼ(electroencephalography, EEG)���Ȼ������Щ�о������������ܵ������������ȣ�������о�δ�ܸ���ͨ��tESע�����������Ϊ���ơ���Σ���һ���о�δ�ܷ�������tES���ڼ�¼��EEG���κ�Ӱ�죬��������ΪtES�շ��ĵ���̫������Ӱ���ڲ�����������(10���ߵ�ǿ��)��Ŀǰ���������ν����Щ���졣���űջ��̼��ķ����� tES����λ���ڲ�����λͬ�������ܻ�����Щ���⡣������Ϊ�ջ��̼�ʹ����tES���������������Ч����ǿ��˯���ڼ���������
WMǿ�������ڴ�������֮���Э������ã��������鵼���ر���theta������Ϊ��WM�з����˹ؼ����ã�����������֯��һ��˳����룬�Ӷ�ά��WM����Ŀ(items)֮���ʱ���ϵ��������鳤���һ���о�������ǰ��Ͷ�Ҷ����֮���thetaͬ��������Ч�ض���WM�е���Ϣ���ƺ����Եó������Ľ��ۣ��ڶ�Ҷ��ǰ��Ҷ��������theta����������WM�ı��֡���ʵ�ϣ�Albouy����ͬ���ṩ��֧����һ�����֤�ݣ�����ʹ��5 Hz��rTMS�̼���ඥ�ڹ�(intraparietal sulcus, IPS)������������theta�̼�����Ҫά����Ŀ����˳��������������WM����(ͼ2A)��ͬʱ��EEG��¼����ʾ��5 Hz��rTMS�����ڴ̼��ڼ���ǿ��theta������������Щ�շ������ڴ̼�������������ڡ���һ������ṩ���ر�������֤�ݣ�֤��rTMSȷʵӰ�����ڲ����������������Ա���Ϊ�д��ز�(��1)������֤����5 Hz�Ķ�Ҷ����̼���������ǰ��ҶƤ���thetaƵ�ʹ������ӣ��Ӷ���ʾ�˾ֲ��̼��ԶҶtheta�����Ӱ�졣���������̼�(transcranial alternating current stimulation, tACS)�о���ͼͨ���̼�ǰ��Ͷ�Ҷ����ʹtheta��λ����(������λ����)������෴(��180����)��ֱ�Ӳ�������ǰ��Ҷ�붥Ҷtheta���ӵ��������ԡ���ʵ�ϣ��������о������֣������첽��(��180����λ�ͺ�)�̼���ȣ�����ͬ����(������λ�ͺ�)�̼��ڼ䣬WM���ֵõ�����ߡ����⣬�ݱ�����ͬ��(����λ�ͺ�)�̼������˶�Ҷ�Ͷ�Ҷ����֮��Ĺ�������(ͨ�����ܴŹ������(functional magnetic resonance imaging, fMRI)����)����Щ�о���һ����Ҫ������ѡ��ĵ缫��̫��(montage)��������Ǵ��ŵģ�������������������������(���磬��ͬ�����ͬ�������¿��ܴ̼���ͬ�Ĵ�������)��δ�����о�Ӧ��ʹ�ø��Ż��ķ��������ܴ�����Щ���⣬�����������о�ȷʵ�������Ҷtheta������WM��Ŀ��ά���о���������á�
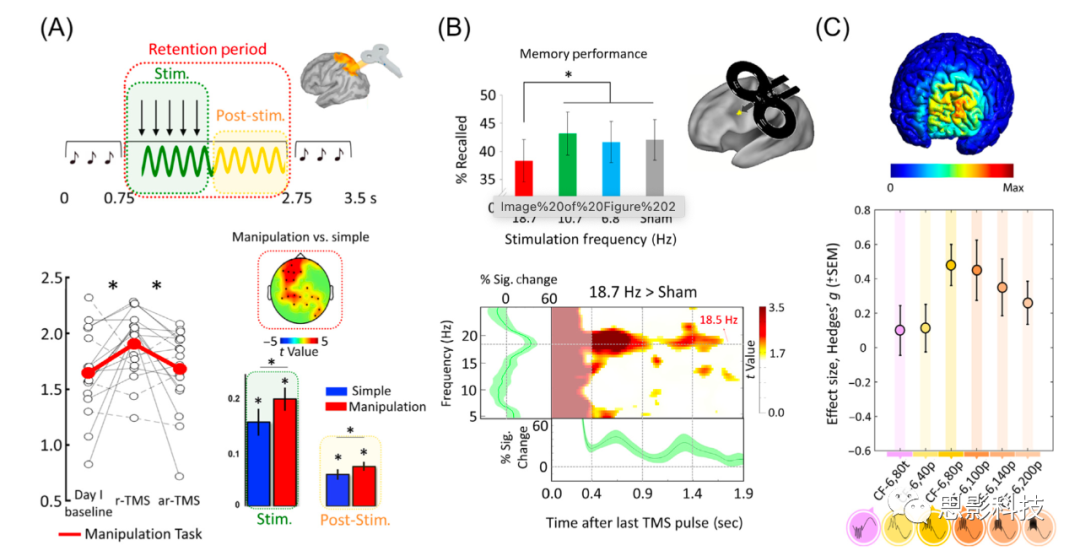
ͼ2 ͨ���������Ե�/�żд������ļ���ЧӦ��
(A)����ߺͽ����Ծ��Ŵ̼�(arhythmic transcranial magnetic stimulation, ar-TMS)��ȣ������������(working memory, WM)�����д̼���ඥҶ�ڹ��������֪���֡����Ŵ̼�(transcranial magnetic stimulation, TMS)����������Ե�ͼ(electroencephalography, EEG)��Ӧ�ȴ̼������ӳ�Լ5������(����)��
(B)��betaƵ�ʴ̼������»ػ�ѡ���Ե������Բ���(����)�ļ�����롣EEG������ʾ���Դ̼�Ƶ�ʣ�18.7 Hz���Ĵ���������Ӧ�ȴ̼�ʱ�䳤Լ1.5�롣
(C)ͨ�������ǰ��ҶƤ��(left dorsolateral prefrontal cortex, DLPFC)��������̼�(transcranial alternating current stimulation, tACS)��theta-gamma����Ƶ����ϴ̼�����WM���֡���80 Hz����theta��(����ɫ)����ϣ�������theta��(��ɫ)�����ʱ��WM���ֵõ������ơ�
����ͨ��Ƕ��gamma����ʱ�Ե���֯WM�е���Ϣ�������˵������gamma���ڿ��ԶԵ�����Ŀ���б��롣Ȼ�����ͨ��Ƕ����thetaѭ���ڵĶ��gammaѭ������Ŀ���н��б��롣��һ����Ԥ�⣬�ⲿ��Ӧ��gamma����theta���ڵIJ���ȿ��ܻͬ�̶ȵ�Ӱ��WM������Alekseichuk������ͬ��ͨ����һ���ռ�WM�������ø��ӵ�theta-gamma���̼���ǰ��ҶƤ��(prefrontal cortex, PFC)����֤������衣Ƕ����theta�����е�gamma�̼������WM���֣���Ƕ����theta�����е�gamma�̼�û�����WM����(ͼ2C)����Ȥ���ǣ�gamma�̼���theta��������ϻ������Գ��ڼ�����롣��theta-gamma WMģ�͵ó�����һ��Ԥ���ǣ�����theta��Ƶ�����������gamma����Ƕ�ף���Ӧ�û�����WM�������෴������thetaƵ��Ӧ�û����gammaѭ�����������Ӷ�����WM���������������tACS�о�֤ʵ����һԤ�⣬�����ڽϵ͵�thetaƵ���´̼���������WM������Wolinski������ͬ�½�һ��֤���������theta��Ƶ�ʴ̼��ή��WM�������������о���ͬ�ṩ��gamma����theta������Ƕ�����֤�ݣ��������һ������WM�п���ά�ֵ���Ŀ������
alpha��beta���������֪��������ѧ�����йأ�һ��ͻ���Ĺ۵���Ϊ���Ƿ�ӳ��Ƥ������Ĺ��������ơ���ˣ����漰�Ӿ��̼���WM�����У���WMά�ֹ�����alpha�����ӱ�����Ϊ��ӳ���Ӿ��ӹ�����Ĺ��������ơ����ֹ������ƿ���ͨ�����DZ�ڵ��Ӿ�������Ϣ�ļӹ���������Ϣ���ڲ�ά�֡�һ��rTMS�о�֧����alpha������������ã����о���������10 Hzʱ�̼���Ҫά����Ϣ��ͬ�ඥҶ����������WM���֣���ͬ���Ĵ̼��Բ�������ή��WM���֡����Ƶ�֤��������һ��tACS�о��������ڶ�Ҷ10 Hz�̼�ʱ�������˵�WM�����������ơ������龰���䣬alpha��beta�ļ���������γ��йء����������Բ��ϵļ����γ�������»ص�beta�����½��йء�rTMS�о�Ϊ���������ϵ�ṩ��֤�ݣ����о���ʾ����betaƵ��(~18.5 Hz��ͼ2B)�������ǰ��Ƥ��ͬ���������������γɡ����߽�һ���������ڴ̼�ֹͣ�д���beta����Լ1.5 s�����仰˵����һ���д�����������������ɸ���Ƶ���Ƿ���̼�Ƶ����ƥ�������ڵģ�������ڲ���beta�������ɴ̼������ġ����������������о�֧��alpha��beta����WM�ͳ�ʱ�����е�������á�Ȼ������theta��gamma���෴���ڻ���������������Ϣ�������У�alpha��beta��ȥͬ���ƺ��Լ������档���ڼٶ���alpha��beta�����ƹ��ܣ�alpha��beta�ļд����ڳ�Ĭ����(���ܸ��ż���ӹ�������)ʱ���������ڼ��䡣����δ�����о���˵��һ����Ҫ�Ŀ��������ǣ��̼�alpha��theta�Ƿ��Լ�����ֲ�����ͬ��Ӱ�죬�����Ӿ�������ʽ����ʾ��gamma��beta��һ����
4. ͨ������̼�(Deep Brain Stimulation, DBS)�������Ե�д�
�������������������йص��о��У�ʹ����ģʽ�������Դ̼��������ӡ���ȻһЩ�о�Ӧ����40 Hz�����Ҳ�(�ڱ�Ƥ��ͺ���֮��)���������о�ʹ���˵�Ƶ�̼�(5 Hz)��theta������̼�(theta-bursts������û�д̼��������ڣ���theta����Χ���Ը�Ƶ�н���ؽ���Ӧ�ü��ִ̼�)��Theta-burst�̼����շ����ݶ���LTP��һ�ַdz���Ч�Ĵ̼����������ҿ�������Ϊģ�������Ϸ�����λ�����ϵ�EEGģʽ���ڼ�ƪ�����У�burst�̼����������ص��ʹ�������������theta-burst�̼������磬Miller����ͬ���ڻ������һϵ��������ѧ����ʱ����һ��������ж��������������theta-burst�̼������Ƿ��֣��������theta-burst�̼����Ӿ��ռ�ѧϰ�����м�ʱ���ӳټ�����ֵĸ����йء�����һ��ʹ��theta-burst�̼���ǿ������о��У�Titiz������ͬ��ͨ������Ƥ��ͺ���֮��Ĵ���ͨ·��ֱ��Ϊ100 ��m�ĵ缫�������̼������Ҳ�������ʵ�theta-burst�̼�����������Ф��ļ��������ԡ�ֵ��ע����ǣ�����ȱ��������������Щ���ڽ����Դ̼����ڼ�����о�����ܵ������ɣ���Ϊ�����ռ����Ե�����������൱�����Ļ���Ⱥ�塣������о��ռ���DBS���ڼ���Ĵ�������������ݼ�(>200������)���������USA�˸����������ɵ�������4���ڼ��ռ�������[�������о��ƻ���(Defense Advanced Research Projects Agency, DARPA)�ָ�����������Ŀ]������Щ�о���Ŀ�IJ�����ͨ���н����DBS���շ��ض�����������ڵ�ǰ����������ļд��Ķ��壬����һЩ�о�������theta-burst DBS������и��ƣ�������Ϊֹ�������о������˶�theta-burst DBS��Ӧ��ʵ���仯��
��������ڼ�����ǿ��DBS�о����������̼���������ı仯����Ϊ�̼���λ���ڵ�αӰ����Ϊ��������д�ЧӦ����һ�ַ�����һЩ�����Դ̼��о������˴̼�ǰ����ı仯��Ȼ������Щ�о������������ڻ��ڴ̼�����Ƶ�ʵ��д������磬Kucewicz������ͬ�±������������ڵ��ʳ��ֹ����дӴ̼�ǰ���̼�����뵥������ʱ��50 Hz�Ĵ̼����Ե��ڸ�gamma (62-118 Hz)�Ļ�������ֿ�Ƶ���������Ӳ����ܾ��巴ӳ50 Hz�̼��ļд��������о�Ҳ̽�����ܴ̼���δ�ܴ̼���Ŀ������̼��ں��һ��ʱ���ڵ��仯����������֪������Ϊֹֻ�������о������˰����ڼ��������ع�����Ĵ̼����������о��˴̼��ں�ļд���ʽ��
Kim��ͬ��ʹ�û���������Դ̼�������ѡ��̼��е㣬��ͼ��ʱ�ռ������ͼ�����ʽ�е��ڼ��䡣������˵�����ǻ��ڵ�������ijɶ���λһ���Զ�����ͼ�������Զ���(���ڵ��)��ֱ�Ӵ̼�������������Ŧ(hubs)���Բ���theta��λ����ڼ�������еı�Ҫ�Ժ�ѡ���ԡ���һ��Ԥ�̼������У�����ȷ���������ռ䡱��ʱ�䡱���������±��ֳ�ǿtheta��λ��ɵ�������Ŧ��������Ϊ�ϣ����Ƿ��ֶ���������ڵ��theta-burst�̼�(4��5 mA, 1����4��50 Hz������̼�)���˿ռ����������Ӱ��ʱ����������Ƿ��֣��ڴ̼�������400�����ڣ����������theta����λ����Կ�ʼ���ӣ����ڴ̼���500�������ң��������翪ʼ�����Щ���ֱ������������Դ̼��������ļд�֮����ڸ�Ϊ���ӵĹ�ϵ���ڴ̼���ֹ�ã�����ļд��ͻ���λ�ڽ�����
Inman������ͬ�²����˶Ի���������ʺ˵�theta-burst�̼��Ƿ������ǿ�����������ʶ����䡣�ڸ÷�ʽ�У��ڱ�������г����������塣���ѡȡһ������������������ʺ˴̼�(��0.5 mA�½���1��8��50����bursts�Ĵ̼���ͼ3)���ڱ�����������ӳ�1�����ÿ�������ʶ����䡣���ʺ˴̼�ȷʵ���������ӳ�1����Եĺ�������ʶ����䡣��Ȼ�̼�αӰ�ų����κζ����ʺ˴̼��������д��ķ����������߲������ڲ��Ҷ(�����ʺˡ�����Ƥ�㡢ǰ����)��ȷʶ����ǰ�ܴ̼��������δ�ܴ̼�������ʱ�ĵ�������Ƿ��пɿ��ı仯(ͼ3A-C)����Ȥ���ǣ����Ƿ�������30-55Hz��gamma�������ӣ������������ʺ�6-8����theta������ض���λ��(����λ�����ϣ�ͼ3D-F������Щ���ֱ������̼������ʺ�-������-�������������ʱ�д���һ�ֱ�theta���Ƶ�gammaģʽ������һ��ʱ���ͻ�������Ժ��̺�����ȷʶ����ǰ�ܴ̼����������塣���������������о��������ڵ�̼���һ�����õ��ֶΣ����յ������ر���theta��gamma���Ӷ�����Եص��ڼ�����֡���Щ�о�֤��������������Ҫʹ���ܼ���ӹ��ڼ��ض��������������������������Դ̼�������������ʹ�÷����Դ̼�����(��50��130���ȵĴ̼�)�����ǵ���һ�㣬�����֤�ݱ�������ʹ�Ƿ��̼�Ҳ���ڴ��Ե����������շ�����������ܵ���˵��δ���������Դ̼��������о�Ӧ���ṩ����������������Դ������Ƶ����ƥ��Ĵ̼�Ƶ��ģʽ���������Щ��Դ�ԡ���Ϊ���Ƶ�ʴ̼��µ�����Ϊ�����
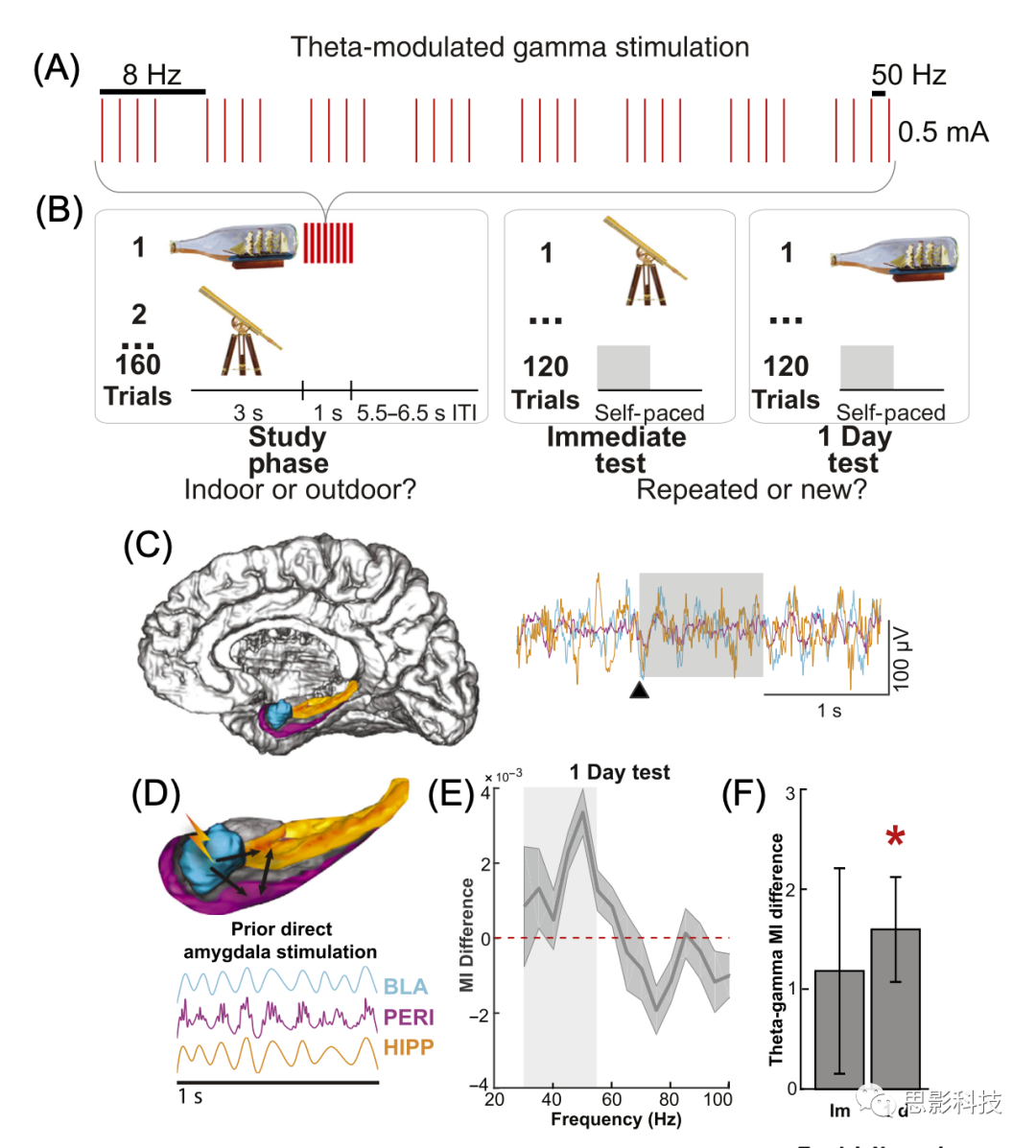
ͼ3 ���ʺ���Դ̼�(deep brain stimulation, DBS)�ļд�ЧӦ��
(A)�������ʺ�1��̼���������ʾ��ͼ(ÿ������Ϊ500 ��s˫���������Ƶ��50 Hz����Ƶ�ʣ�8 Hz)��
(B)ʶ���������ʾ��ͼ�����о�������̼�һ������壬�����о��κ�������1��Զ��ص�ͼ���Ӽ�����ʶ�������ԡ���ɫ��Ӱ�����Ӧ��ʶ������ڼ�ͼƬ���ֺ��ǰ0.5�룬Ҳ�����(C)�Ҳ���ʾ����Ϊѧ������������������ʺ˽��ж��ݵĵ�̼�������ǿ���ij����Լ��䣬����������������Ӧ��
(C)��ʶ����������У��Ի���������ʺ�(basolateral amygdala, BLA)������(hippocampus, HIPP)�ͱ���Ƥ��(perirhinal cortex, PERI)�Լ�ÿ����������Ծֲ�����λ(local field potential, LFP)��ͼʾ(��ɫ�����α�ʾͼ��ʼ;��ɫ��Ӱ�����Ӧ��ʶ�����ʱ���B�е���Ӱ����)��
(D)�̼�������������BLA��HIPP��PERI��1��ʶ����������ʾ��ͼ����Щ������������������֮��theta����õ����ӣ��Լ�����Щ�������ڵı���Ƥ���gamma������
(E)��1������У��̼����̼������±���Ƥ����Ƶ�ʵĵ���ָ��(Modulation index, MI)���졣��Ӱ�����ʾ30-55���ȵ�gamma���Ρ�
(F)�̼�-�̼������±���Ƥ��gamma��Χ���ۻ�MI���졣�ڵ�1��(1 d)�����У��̼������¼���ͼ���MI���̼������¼���ͼ���MI���ӡ�
5.δ���ķ���;�����
�����������о�������ͨ���д������д����������ض�������һ�ֺ���ǰ���ĵ��ڼ�����ֵ�;����Ȼ����ֵ��ע����ǣ�����������о���������Ҫ�ķ���ѧ�����ԡ��ر��ǣ����������Ĵ�����о������������Լ����о���ÿ��ʵ��ʹ�����൱�͵�������(��1)������������ڷ���ƫ�кͼ����Է��ն��߹�ЧӦ�������⡣�����һ��tDCS�о�ʹ�����൱���������(��75��������)������ЧӦ��Ϊ0.45������ζ�ż�ʹ�������о�Ҳ���в��㡣��ǿ����ʹ���ʵ��Ĵ��ģ�о�����Ҫ�ԣ��Լ����ظ�����������������ƪ�������������ֵı�Ҫ�ԡ����⣬����ϣ����δ����������ʹ��Ԥע��Ĵ̼��о����⽫���ӽ�������ȺͿɸ����ԡ�
����ڴٽ���������д�����Ļ����Լ�������ξ����Ӱ�����Ҳ����Ҫ��Ϊ�ˣ�������Ҫ�������ռ���Ϊ���ݵ�ͬʱ�ռ������������ݣ��Ը��õ��˽�Ŀ�����Ƿ��Լ�����ܵ��д���Ӱ��(��1)����ʵ�ϣ�����ع˵Ĵ�����о���ʹ����ij����ʽ���������������Խ����Դ̼��Ƿ�Ӱ����Ȼ������Ҫ���������õķ�����ʹ���ܹ��ڴ̼��ڼ������пɿ��IJ������������ڴ̼�αӰ�����⣬Խ��Խ���Ե��ǣ��̼�����Ӧ�ø����ڲ����綯̬���е���������������ȷ�����ر������Ŀ�������(�����Ǹ���)������֡�ֱ�۵�˵��������ķ����Ƚ����Ե�������Դ�Թ��̵ķ�����ö࣬����Խ���Ϊʲô��ǿ����Ĵ̼������ر������ս�ԡ��ջ��̼��IJ������ã������д��̼�����λ��Ƶ�ʺͲ������ڲ�����ѧ��ƥ�䣬�Լ��������������Ĵ̼����������ⷽ������ǰ;�ķ�չ����
�����Դ̼���������Դ̼��IJ�ͬ����(���磬���ڼ�¼���Ŀռ�ֱ��ʺʹ̼�ЧӦ�ľ�����)���ܲ���������Ҫ�IJ�ͬ�������Щ���Ӧ����δ������ϵͳ�ıȽϡ���Ȼ�д��̼��ڰ����ض���������ʱ���ܸ���Ч�������̼���ȫ��ЧӦ���ܵ��¶Զ���������ϵͳ�ĸ���ķֲ�ʽЧӦ����ˣ�δ�����о���Ӧ���Խ������Լ�¼������ÿ�ִ̼�������ԣ��Բ�����Э���������Դ̼��������Դ̼��Ļ��ƻ��������Ż�������ǿЧ�������磬�п��ܶԾֲ����ƣ������Ƕ�ȫ�ּд�����ʵ�ּ�����ǿ�Ĺؼ���δ�����о���Ӧ����д���¼�����̼�(��tES)���Ը��õ������ȫ�ֵ����д�ЧӦ���Ӱ��ֲ������⣬�ڱջ��̼��У��̼�������ʵʱ��������Ӧ�������п����ڼ����з��벻ͬ�����ơ�������ڻ����Խ��д��̼�������¼�������ϣ��Ӷ�����㷺����ЧӦ����ЩЧӦ�ڽ��������������и�Ϊ�ձ�������о���
������Ҫ���õ�����д���ϸ����·�ĸ���������ѧӰ�졣Ϊ�˽����һ���⣬��Ҫ���и����о�����������λ��¼���Ŵ��̼�(�ڶ���ģ����)��/���̼�(���������ģ����)���ϡ����������罨ģ����Ҳ���DZز����ٵģ���Ϊ�������ṩ�ɲ��Ե�Ԥ�⣬���������ڵ�����λ�ͺ��ˮƽ�ľ�����֮��ת����
�ܽ
��������������ǣ������Ƿ�������������ϵ�����Ǹ�����һ�ָ������������������о�ʹ���˸�����ʽ�ļд���֧����ǰһ�ֹ۵㣬��������ʵ����ʵʩ���ض������ƣ�����������γɡ�ά�֡����̺���ȡ�������ڲ����д��������Ӱ����䷽�滹�кܶ��Ҫ�����������ڿ��Կ�ʼ��������ڼ��������ʵʩ�ľ�����ơ�����ڿ���������ؼ�������Ч���Ʒ���������Ҫ��
����ԭ�ļ��������������˼Ӱ�Ƽ��ţ�siyingyxf��18983979082��ȡ,���˼Ӱ�γ̼��������ȤҲ�ɼӴ��ź���ѯ����˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ�����������ǵĽ���������о��а����������ת��֧���Լ����½ǵ��һ���ڿ����Ƕ�˼Ӱ�Ƽ���֧�֣���л��

��ɨ����߳���ѡ��ʶ���ע˼Ӱ�dz���лת��֧�����Ƽ�
��ӭ���˼Ӱ�����ݴ���ҵ�γ̽��ܡ�����ֱ�ӵ���������ּ������˼Ӱ�Ƽ����еĿγ̣���ӭ�����ź�siyingyxf��18983979082������ѯ�����пγ̾����ű��������������ǻ��һʱ����ϵ���������ѱ���ѧԱ�����
�Ե缰���⡢�۶���
������
����ʮ�����Ե����ݴ����м��ࣨ������4.7-11��
�ڶ�ʮ����Ե����ݴ������Ű�(������5.16-20��
�Ͼ���
�ڶ�ʮ�����Ե����ݴ������Űࣨ�Ͼ���4.23-27��
�Ϻ���
�������Ե����ѧϰ���ݴ����ࣨMatlab�棬�Ϻ���5.15-20��
����ʮ�����Ե����ݴ����м���(�Ϻ���5.28-6.1��
���죺
�ڶ�ʮ����������Թ������ݴ����ࣨ���죬5.24-29��
�˴ţ�
�Ͼ���
�ڶ�ʮ�Ľ�DTI�ࣨ�Ͼ���4.9-14��
������
����ʮ�Ž�Ź�����Ӱ������ࣨ������4.14-19��
�ڶ�ʮ�Ž�Ź������������ݴ����ࣨ������4.22-27��
�ڶ�ʮ���DTI�ࣨ������5.6-11��
��ʮһ��Ź���ASL�ࣨ������5.12-15��
���죺
��ʮ�Ž�Ź�����Ӱ��ṹ�ࣨ���죬4.13-18��
����ʮ��Ź�����Ӱ������ࣨ����,4.21-26��
�����Ӱ����ѧ�ࣨ���죬5.19-23��
�Ϻ���
���Ľ�Ź���Ӱ����ѧ�ࣨ�Ϻ���4.13-17��
��ʮ��������̬���ܴŹ������ݴ����ࣨ�Ϻ���4.21-25��
����ʮһ��Ź�����Ӱ�������(�Ϻ���5.7-12��
���£��ڶ�ʮ�˽�Ź������������ݴ����ࣨ�Ϻ���5.22-27��
���ݴ���ҵ����ܣ�
˼Ӱ�Ƽ����ܴŹ���(fMRI)���ݴ���ҵ��
˼Ӱ�Ƽ���ɢ��Ȩ����DWI/dMRI�����ݴ���
˼Ӱ�Ƽ��Խṹ�Ź����������ݴ���ҵ����T1)
˼Ӱ�Ƽ������ද���С����Ӱ�����ݴ���ҵ��
˼Ӱ���ݴ���ҵ������ASL���ݴ���
˼Ӱ�Ƽ���Ӱ�����ѧϰ���ݴ���ҵ�����
˼Ӱ�Ƽ������Ⱥ����ҵ��
˼Ӱ�Ƽ�EEG/ERP���ݴ���ҵ��
˼Ӱ�Ƽ��������Թ������ݴ�������
˼Ӱ�Ƽ��Ե����ѧϰ���ݴ���ҵ��
˼Ӱ���ݴ������������Դ�ͼ��MEG�����ݴ���
˼Ӱ�Ƽ��۶����ݴ�������
��Ƹ����Ʒ��
˼Ӱ�Ƽ���Ƹ���ݴ�������ʦ ���Ϻ����������Ͼ���
BIOSEMI�Ե�ϵͳ����
Ŀ��ʽ���ܴŹ���̼�ϵͳ����