���Թ���ҽѧԺ��Tang wei��Stufflebeam������PNAS�Ϸ��ģ���Ҫ����������Ծ�Ϣ̬�����������Ķ�̬����Ӱ��ֲ�������о�������ָ������Ŀǰ��Դ���ģ��ĸ���;ۺ��д������о������Ƕ��ڷ����ģ��֮����ν��оۺϵ��˽���Ȼ���㡣�������ĺ�����̽Ѱ�����������������Ӱ����仯�Ļ��ơ�����������о�̽Ѱ�������������������Ӱ���ض����������ͨ������ͬһ�鱻�ԣ�29���������ľ�Ϣ̬fMRI�Լ�MEG����,����������Ϣ̬���磨DMN��������������ľ�Ϣ̬��ϵ���õ�10-Hz��Χ�ڣ���۴�Ƥ�ʣ�PCC����ǰ�۴�Ƥ�ʣ�ACC���Ķ���Ӱ�졣������ʱ��仯��Ӱ����ACC�źŵ�ǿ�ȱ仯��أ���13-16HzʹƵ��ǿ���½���1-7Hz��30-55Hz��ʹƵ��ǿ�����ӡ��о������֣�30-55Hz�������ACC��3-4Hz�ļ�����λ��ϡ���Щ��������ֲ�Ƶ�ױ仯��������֮���������ԡ���ЩƵ����Ϣ���Ա���PCC-ACC�����ھ�Ϣ̬�µĹ��������ã�ͬʱ��������ֲ�����ʹ������綯��ѧ֮�����̬��ϵ��
��߶��Ի��һ�����������Ǹ���;ۺϡ���ͬ��������������֪�����б�������γ�һ�������Щ���������ͨ�������������Ӱ�죬���յ���ȷ����Ϊ�������ۺϽ��������;ۺ���������ż����ȷ������֪�����е�������Ҫ�о�����1����Ӧ��֪�ӹ��̵������ֲ�������о�;2����������ϵ��о����䷴Ӧ��ģ��֮��ľۺϡ���Ȼ��Ӱ��ѧ���������������ϵ��о����н�չ�����о����֮���Դ��ڲ�ࡣ���ڸ���ģ�����ξۺϵ�����ѧ�������˽���Ȼ���㡣������ĺ������˽�һ����������������Ӱ���µľֲ�������Ӧ���ƣ�һ����Ҫ�IJ����ǵõ��������һ�������Ȼ����Ŀǰ���о���Ҫ������������ļ���������������ӵ��������о�����û�н�һ����������֮��Ĺ�ϵ����̽���������о��������������ͬʱ�о�������ͨ�Ժ�����������ַ�ʽ��Ҫ���кܸ�ʱ�շֱ��ʵ����ݣ������Ŀǰ�ij����������Ǻ���ʵ�ֵģ��������ʼ���о���ͼ�ֲ���Щ��ࡣһ�ַ�ʽ�Ƕ�fMRI�����û�������ʽ�������Ӷ�����ʱ����Ϣ������һ�ַ�ʽ������MEG������fMRI��ϣ���֤������ͨ�Ժ�������Ĺ�ϵ����Щ�·����������������ʱ�չ�ϵ���Ӷ��ܹ����������Ӻ����������ϵ�����о������ǵ���Ϣ̬���磨DMN���ھ�Ϣ״̬�¿��Ա���̼���������ļ�����ţ��Լ���DMN�Ƕ��ڳ������о���֪���ܷ����dz���Ч����ˣ��о�ѡ�����о�DMN���������������Ե��о����о��ɼ���ͬһ�������ߵ�fMRI��MEG���ݣ���������������DMN��������������ǰ�ͺ�۴�Ƥ�ʣ�ACC��PCC������fMRI��ָ������MEG������ACC��PCC�Ķ�ʱ�����������ϵ��GC�������ֶ�̬�����ܹ������Ǽ����������������Ĺ��ױ仯�Լ����ֱ仯������������������صĹ�ϵ���о������������֮���������ܹ�ʵʱ��Ӱ�������ڼ���ر�أ��о�ָ��PCC��һ��Ƶ���е�Ӱ����ACC�ڶ��Ƶ���еľֲ�Ƶ�ױ仯��ء����⣬�����ڲ�Ƶ����������λ���ȹ�ϵ����ʾ��PCC-ACC֮��Ĺ�����ϵ��
��
����
���ȣ�������֤��MEG�ռ��л�õ�ACC-PCC���ӵ�GC�ɿ��ԡ�������̰����������裺�������ù�������MRI��fcMRI����Ϊָ��������MEGƵ���е�ACC-PCC���ӡ��������ø�Ƶ�����ACC��PCC֮��ĵ�GCͼ�����ţ��о���֤��GC��ʱ��仯�Ƿ���MEGԴ�źŵľֲ����ʱ仯��ء��Ƚ���GC��һ�������ǿ��ʱ��κ͵�GC����ʱ�Ĺ������ܶȣ�PSD���� ������ʾPSD�����Դ�źŵ�Ƶ�ʷ������о������˽���Ƶ���������֤���Ե�ģ�鼤���Ƿ����һ���Ĺ������塣����ѡ��10 Hz�������fMRI�����ӡ�����MEG֮�书�ʼ���ACC-PCC֮���ʱ������ԣ���ΪMEG��ͨ�ԵIJ�ȡ���������������Դ�źŵ����ͺ���ء�����1��56Hz֮���ACC-PCC��ͬƵ�ʵĵ�����ԡ�����10Hzǿ�ȵ��������ߣ�ͼ1A�������⣬�仹����BOLD�źż�����ACC��PCC֮���fcMRI��10Hz���ʵõ������Ӻ�fMRI���Ӿ�������������������ԣ���Pearson��s r = 0.39, P < 0.04, ͼ1B������ʾ����10Hz�����¸�����ǿ��fcMRIͨ������ǿ��MEG���ӡ���ˣ��о�����10Hz�����µ�����������ACC��PCC֮������ӡ�
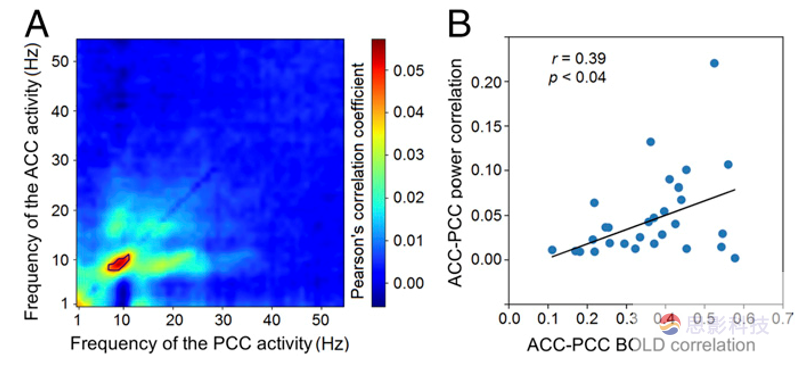
ͼ����MEG������10Hz����fcMRI�����к�ǿ������ԡ���A��ACC��PCC��MEG�źŵ�ʱ������Ե�ɢ��ͼ����ɫ����Pearson��صĴ�С�����ұ��������rֵ����B��FcMRI��10Hz���ҵ�MEG����ɢ��ͼ��ÿ���������������ɫΪ�ع��ߡ�
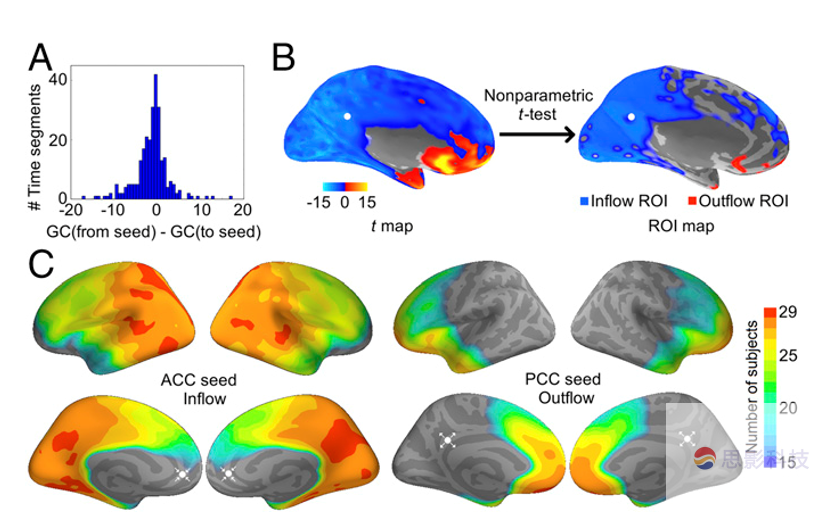
ͼ����10Hz�µ�GCͼ��һ���ض�������ʾ������ʾΪ���ӵ�������������һ��ʱ����ڵ�GCֱ��ͼ,ֵΪ���ʾû��GC����B��PCC���ӵ�������Լ��顣��C��ACC�����루��PCC���������ң����鼶���ơ�
�������ӵ��ACC��PCC֮��Ŀռ�������GC����
���û�����������ÿ��������10Hz��Χ�ڵ�ACC-PCC��GC����Ϊ�������Ķ�̬MEG���Ӳ�ȡ��ڲ�����ͨ�Ժ������֮��Ĺ�����֮ǰ�����ȼ��GC�����Ŀռ������ԡ���һ��ͬʱ����֤GC��Ч����֤����ͨ�ԣ��Լ��ų�������ǰ����������ACC��PCC�����Ԥѡ�����DZ��ƫ�Ϊ�ˣ�ʹ�û������ӵ�ķ�������һ������������Ϊ����ʱ��GC����Ӧ��������������Χ��ʾ��ľ���ֵ������Ƥ������ಿ����ʾ�����Сֵ��������ʹ�ó����ӵ�������ӵ��GC֮��IJ�ֵ��ͼ2A���������ԭʼGCֵ�����ֱ����Ϊ�������������ֱ��Ϊ������������ͬʱ��ν��зDz�������ˮƽ���ԣ���ȷ��ÿ�������GC���������ԡ���Ч���㱻���Ϊ����Ȥ����ROI������ʾGCֵ�ϴ������ͼ2B�����õ�ROI����ÿ���������л���ROI��Ȼ���ڶ���֮�串���Եó��鼶��Ĺ��ƣ�ͼ2C��������ACC�������������Ӱ�죬������29���������У�PPC���ֳ����ڶ��յ�GCӰ�졣����PCC�������������Ӱ�졡����28���������У���GCӰ�죬ACC��ΪROI��������ACC����ͼ��ACC�� ROI�е�PCC����ͼ�е�û���ҵ�PCC�� ROI��ͼS1��S2�������һ�µر�����GCӰ������PCCֱ��ָ��ACC�ġ�
ACC����AC��صĹ����ױ仯
�������ӵ�GCͼ������֤��ACC-PCC����õĿռ������ԣ�������������õķ��Գ��ԡ���PCCָ��ACC�и�ǿ��GC����֮��Ȼ���о�̽Ѱ��ACC�еľֲ����θ���PCC��GCӰ����ı䡣������˵���о������˲�ͬGCǿ����ACC��PSD��ǿ�ȡ���ÿ��ʱ�䴰�ڣ�����ACCÿ�������GC��PSD��ͼ3A�������������ʱ��ξ����ʱ�մأ��õ�ʱ�䣨> 20 s���Ϳռ��ȶ���> 20�����ڶ��㣩��ʱ��ģʽ�����ݶ�������ݶ��Ƿ�����GC�أ���Щ�����ױ����Ϊ�ڲ��ػ��ⲿ�ء����ڵ����ݶ���ʾ����13-16Hz��Χ�ڵĹ���ʼ�սϵͣ����� 1-7Hz��30-55Hz�Ĺ��ʷ�Χ�ڵĹ��ʸ���ͼ3B�������в���������б������������S1����Щ����ṩ�˴�PCC��ACC�Ķ���Ӱ����ACC���ʱ仯֮��Ĺ�ϵ���������ض�����֪��У�һЩ���ļ�����λ��theta������Ӱ��һ����ļ�����λ��gamma������Ϊ�ˣ��о�������Ƶ�Ͷ˺߶�֮���Ƿ���ڽ���Ƶ����ϡ�����ܹ���theta��gamma����֮�䷢�����ģʽ��ͼ3B���������ṩ֤�ݱ������ײ�������ģ����ǻ�Я��һ���Ĺ�����Ϣ������о�������3-4Hz��30-55Hz����֮���ACC���ģʽ��gamma��3- 4-Hz���ȡ���/ 2��λ�ĸ����ﵽ��ֵ��ͼ3C���� ��������֤ʵ����һ���۵������ԣ�ͼ3D����������˵�����û����λ��ϣ���gamma��ǰ5�����ȵ���λ�ǽ���[-�У���]�Ϸ��Ӿ��ȷֲ���Ȼ����ʵ�ʷֲ�����ȷֲ�������ƫ��(ͼ3)����������Ҳ��֤���������ԣ���992= 141.1��P <0.01��������1-7-hz�IJ����У�û�з���������Ƶ�ɷֵ��������
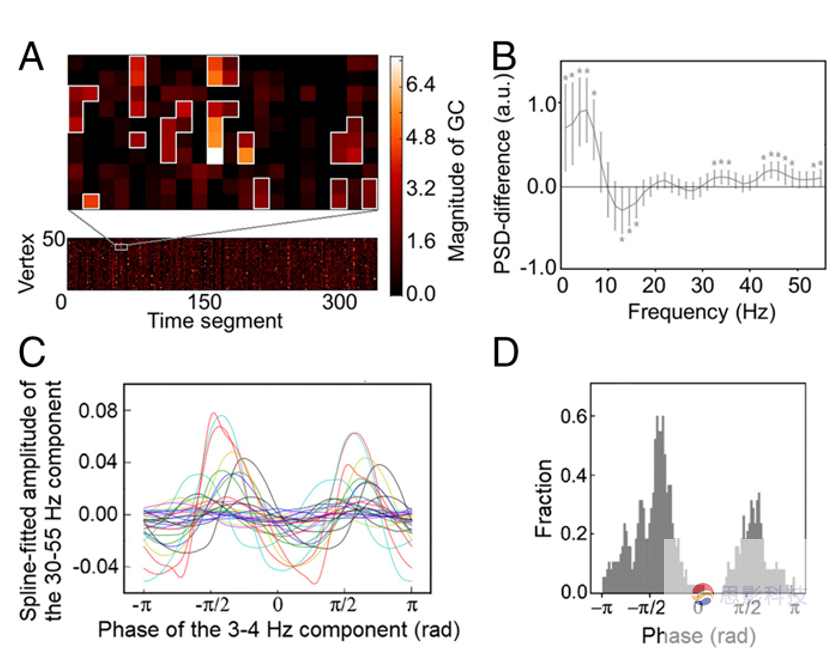
ͼ3 10-Hz�ֲ���仯���GC�����йء�һ�������ʵ������ʾACC��GC�ء���B���������Ƶ���ʲ��죬��ƽ��ֵ��ʾ���Ǻű�ʾ�����ƫ���C��ACC��3-4Hz��λ�ϵ����Gamma�������ʾ��ʾ����/ 2�����ﵽ��ֵ��ÿһ���ߴ���һ��������D����λ��ֱ��ͼ��
����
���½�ʾ��DMN���������������Ķ�̬���Ӻ;ֲ�����Ĺ�ϵ������fcMR��IMEG��������10Hz��PCC��ACC�����ǶԳƵ�GC�����֣���ʱ��仯��GC������ACC��ǿ�ȵı仯���仯��13�C16Hz��Ƶ�������½��Լ�1�C7 Hz��30�C55 Hz��Ƶ�����������ᵼ��ǿ��GC������ACC�У���Ƶ�ɷֺ�Ƶ�ɷֵ����֮��Ҳ������������ϡ���Щ���ֵĴ��Ի�����Ϻͷ���֮�����ϵ����ʾ��DMN�������䶯̬�Ľ������á�
�������Ӻ;ֲ���������������
֮ǰ���о�ͨ���������о��������Ӻ;ֲ����Ȼ��������ƪ�����У����߷���������������ʱ��߶��¾���һ��������ԣ���Ҫָ�����ǣ����ֹ�ϵ�����ܼ����������ԣ��������������ÿ��ܵ���һЩƵ��ǿ�ȵIJ��������⣬�����ľֲ���̬�仯Ҳ����Ӱ��������ϣ�Ŀǰ�ķ����������������ָ��ţ�������ˣ�����о��Ա����������Ĺ��������ṩ��һ�ֽ��͡�
ACC-PCC�Ĺ���������10HzƵ���µ�����
��֮ǰ��һϵ���о��У���Ҫ��һ����Χ��Ƶ���й�����Ϣ̬���硣����alpha��beta���ʹ�õ�����Ƶ�ʷ�Χ�������о���Ҫ����alphaƵ��������ACC-PCC���ӣ�ͼ������ͨ��GC���������ڳ�������Ƶ���µ�PCC��ACC��ֱ�Ӷ�����ϵ��ͼ2C�������⣬��theta��gamma����(ͼ3B)��ACC����GC��������������������������ص�Ƶ��ǿ�Ȳ�������������أ�������ۿ���ͬʱ�ڹ�����غ�fMRI����ϵĵõ���֤������Ľ��Ҳ������֤��������ۣ������⣬������֤������10HZ������õĽ����
���Գ�GC֧��PCC�������hub�ڵ���һ���ۡ�
�о�����PCC��ACC�����GCӰ��ȷ������Ҫǿ��ͼ 2������������������Ϣ����Ҫ��alpha/beta������һ�¡����ֲ��Գ�Ҳ��PCC�ڴ���Ƥ���еĽṹ����hub���ĵ�������һ�¡��ر���һ���о��У�PCC�����ֿ����ڲ�ͬ��״̬����֮����н������������ACC�Ķ���Ӱ���������DMN��ִ�е�PCC��Э�����һ���֡������������ACC����ع��ױ仯���������ڲ�������Э�����������⣬û�й۲쵽��ACC��PCC��GC��GC�е�������λ��һ�ֿ������ǣ������̫�ٵ�theta���ڳ��ȣ�����Ч�Ķ�GC���й��ơ�������ҶƤ�㱻��Ϊ���к�ǿ�ľ�Ϣ״̬alpha��������GC(ͼ2C)�в�û�еõ�һ�µ�ROI����Σ���Ȼ������Ƶ�ʴ����п��ܴ���SNR���죬����������Ƶ�ʵ�GCӳ����û�еõ�һ�µ�ROI(ͼS1��S2)����������ȻƤ������ϵ������MEG����ԴҪ�ߣ������䲢û����ʾ��MEG��GC������ļ�ֵ(ͼS1��S2)��
����Ƶ����Ͻ�ʾACC��Ƥ���˷ܵ����е�����
�������߶���DMN�ĺ���������ACC�Ĺ�����PCC��ͬ���ڻ����������Ϊ�о��У�ACC����Ϊ��һ����ִ�й��ܺ�Ŀ�굼����Ϊ��ص���Ҫ���������theta��gammaƵ��֮�䷢�ֵ��������ϵ����ΪƤ���˷��Ե����������䡣�ھ�Ϣ״̬�£���һ��ִ�й��ܵĹ������ϵ�����ʹ�����ܹ����شӱ���״̬�л���������Ϊ״̬����һ���ֿ���Ϊ��һ���о���һ�����ṩ��һ������ ��
�����������ᵽ������ڵ������ؽڵ�������(��PCC��ACC��Ӱ����ܻᾭ��������������GCͼ����ֱ�Ӽ���õ�������Ŀǰ���е�ӳ�串���˴���Ƥ������ж��㣬������ڣ����ص�Ӱ��ܿ���������Ƥ���µĽṹ������ȱ��Ƥ���µļ�¼�������������Ŀǰ�������еõ������
�������Tang W, Liu H, Douw L, et al. Dynamic connectivity modulates local activity in the core regions of the default-mode network[J]. Proceedings of the National Academy of Sciences, 2017, 114(36): 9713-9718.
��ȡԭ�ģ�������ϼ���������ע��˼Ӱ�Ƽ������ںţ��ظ������·��롱����ȡ�������ӡ�
